
|
Pour le comité de rédaction de la Plateforme ESA : Jean-Philippe Amat, Sophie Carles, Julien Cauchard, Céline Dupuy, Sylvain Falala, Guillaume Gerbier, Viviane Hénaux, Yves Lambert, Renaud Lancelot, Carlène Trévennec
Autres experts sollicités : Mathieu Guillemain1, Sophie Le Bouquin-Leneveu2, Éric Niqueux2, Béatrice Grasland2, Axelle Scoizec2, Audrey Schmitz2, Anne Van De Wiele3
Auteur correspondant : plateforme-esa@anses.fr
- Office Français de la Biodiversité, Unité Avifaune Migratrice, Arles, France
- Laboratoire national de référence de l’influenza aviaire, Anses Ploufragan-Plouzané-Niort, site de Ploufragan,
- Office Français de la Biodiversité, Unité Sanitaire Faune, Auffargis, France
L’encadré 1 précise la fenêtre temporelle, la couverture géographique et les terminologies utilisées au niveau européen pour déclarer les foyers et cas d’IAHP et la notion de pathogénicité des souches d’influenza au sens de l’OIE.
| Encadré 1 : Fenêtre temporelle, couverture géographique et terminologie |
|---|
|
Période Sources de données et couverture géographique Jusqu’au 31/12/2020, les déclarations ADNS (ancien système de déclaration, remplacé par ADIS depuis avril 2021) du Royaume-Uni concernaient l’ensemble du territoire de Grande-Bretagne et d’Irlande du Nord. Depuis le 01/01/2021, ces mêmes déclarations ne concernent plus que le territoire d’Irlande du Nord. Les déclarations du Royaume-Uni traitées dans ce document sont issues de l’OIE/Empres-i pour les figures, et du DEFRA pour le décompte des cas et foyers (Tableau 1), à partir de cette date. Les déclarations à l’OIE/Empres-i sont incluses pour la partie occidentale de la Russie, délimitée par le massif de l’Oural (à l’ouest de la longitude 60°), ainsi que pour l’Albanie. Catégorisation des déclarations à l’ADIS
Classification des virus influenza |
Des premiers foyers et cas sauvages déclarés en Russie occidentale
La Russie a notifié à l’OIE le 04/08/2020 un foyer d’IAHP H5N8 ayant débuté le 27/07/2020 au sein d’un élevage familial (156 oiseaux) dans une région au sud du massif de l’Oural et dans l’ouest de la plaine de Sibérie occidentale, à la frontière kazakhe (source : OIE notification immédiate 04/08/2020). Une carcasse de canard sauvage avait été retrouvée proche du village où se situait l’élevage et testée positive pour H5N8 peu de temps avant la déclaration de ce foyer (source : Commission européenne ADNS au 09/08/2020). La première déclaration à l’ouest du massif de l’Oural (60° longitude) a été reportée le 21/09/2020 (source : OIE le 21/09/2021). La plupart des foyers domestiques détectés ensuite impliquait des basses-cours ou des élevages familiaux. Trois foyers ont concerné des exploitations de type industriel. Pour l’ensemble de ces foyers, un total de 1 557 797 volailles (10 299 animaux morts) a été concerné. Les foyers domestiques étaient situés dans la zone au sud de l’Oural à l’exception de deux foyers détectés à plusieurs centaines de kilomètres à l’ouest (source : OIE rapport de suivi le 05/10/2020). Le 15/10/2020, un total de 55 foyers domestiques et sept cas en avifaune sauvage d’IAHP H5 (incluant la première confirmation du 28/07/2020) avait été déclaré (Figure 1). La majorité a été confirmée de sous type H5N8 de clade 2.3.4.4b de la lignée A/Goose/Guangdong/1/96 (OIE rapport de suivi le 14/10/2020).
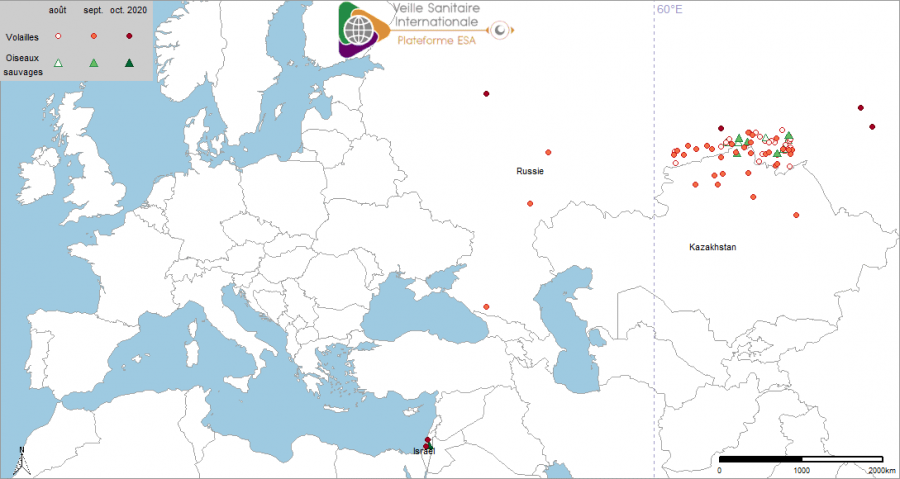
Figure 1 : Localisation des cas en avifaune sauvage et foyers domestiques d’IAHP en Sibérie occidentale, au Kazakhstan[1], au Moyen-Orient[2] et en Europe ayant débuté entre le 01/08/2020 et le 15/10/2020 (source : Commission européenne ADNS, OIE/Empres-i).
Extension rapide en Europe dans les compartiments sauvage et domestique
Les Pays-Bas ont notifié le premier cas en faune sauvage le 21/10/2020 et le premier foyer domestique le 30/10/2020. Plusieurs cas en faune sauvage, puis des foyers domestiques ont également été notifiés au Royaume-Uni et en Allemagne à partir de la fin octobre.
- Aux Pays-Bas à partir du 16/10/2020. Des cas groupés d’IAHP dans l’avifaune sauvage, trois canards siffleurs (Mareca penelope) et un cygne tuberculé (Cygnus olor), ont été détectés le 16/10/2020 et confirmés à partir du 20/10/2020. La première notification est faite le 21/10/2020 (source : Commission européenne ADNS du 21/10/2020 au 04/11/2020). Le premier foyer domestique a été confirmé le 29/10/2020 sur suspicion clinique (mortalité de 1 % au moment de la suspicion) dans un élevage de 35 750 volailles, Gallus reproducteurs en filière chair (source : OIE le 30/10/2020). On constate des délais de notification rapides de ces premiers cas/foyers dès leur confirmation (1 jour).
- Au Royaume-Uni à partir du 24/10/2020. Un foyer domestique ayant débuté le 24/10/2020 dans le comté de Cheshire au nord-ouest de l’Angleterre a été confirmé le 02/11/2020 et notifié le 03/11/2020 (source : Commission européenne le 03/11/2020). La première notification signalant un cas sauvage est datée du 10/11/2020 (source : Commission européenne ADNS le 10/11/2020), mais les premiers cas identifiés au sein de l’avifaune sauvage ont débuté le 26/10/2020 dans le comté de Lancashire, voisin du Cheshire (source : Commission européenne ADNS le 15/12/2020).
- En Allemagne à partir du 26/10/2020. Le pays a déclaré le 02/11/2020 trois premiers cas dans l’avifaune sauvage, ayant débuté le 26/10/2020 et confirmés le 30/10/2020 dans le nord du pays, dont deux canards siffleurs trouvés morts sur des lieux distincts et le troisième chez une buse variable (Buteo buteo) trouvée morte et infectée par un virus appartenant au sous-type H5N5 (source : Commission européenne ADNS au 02/11/2020, LRUE). Les premiers foyers domestiques ont été confirmés le 04/11/2020 dans le nord du pays. Le 09/11/2020, le pays comptait 99 cas sauvages et deux foyers domestiques.
Quatre semaines après la première confirmation aux Pays-Bas, 369 cas sauvages et 21 foyers domestiques ont été notifiés dans dix pays (Allemagne, Belgique, Croatie, Danemark, France, Irlande, Italie, Pays-Bas, Royaume-Uni et Suède) (Figure 2). A ce moment-là de l’épizootie, trois sous-types ont été rapportés : H5N1, H5N5 et H5N8. Le sous-type H5N8 était majoritaire.
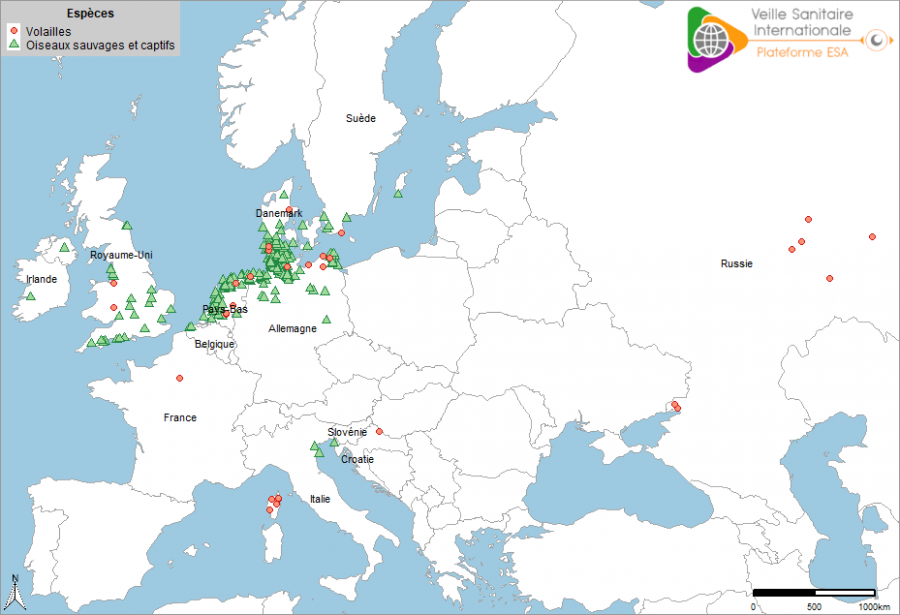
Figure 2 : Localisation des cas en avifaune sauvage et foyers domestiques d’IAHP en Europe ayant débuté pendant les quatre semaines (du 16/10/2020 au 17/11/2020) qui ont suivi la confirmation du premier cas en Europe occidentale (source : Commission européenne ADNS, OIE).
Epizootie en vagues successives
Entre le 01/08/2020 et le 31/07/2021, 2 659 cas en avifaune sauvage (libre ou captive) et 1 303 foyers domestiques ont été confirmés, dans 29 pays (Tableau 1). Trois pays ont totalisé plus de 80 % des foyers domestiques : la France (n=492, dont 475 localisés dans le Sud-Ouest), la Pologne (n=357) et l’Allemagne (n= 235).
L’Allemagne a regroupé près de la moitié des cas sauvages (n=1 332). De nombreux cas sauvages ont également été déclarés dans les pays du nord de l’Europe : Danemark (n=318), Royaume-Uni (n=317), Pays-Bas (n=114) et Suède (n=109).
Tableau 1. Nombre de foyers chez les volailles et de cas sauvages d’IAHP H5 confirmés en Europe par pays du 01/08/2020 au 31/07/2021. Le “x” de H5Nx signifie que le typage de la neuraminidase est inconnu (Source : Commission européenne ADIS pour l’Europe, DEFRA pour le Royaume Uni, Empres-i pour l’Albanie, OIE pour la Russie occidentale au 31/07/2021).
| Pays | Compartiment | Date de première confirmation | Nombre de foyers et cas confirmés pour la saison 2020/2021 | Sous-types identifiés pour l’ensemble des foyers et cas de la saison 2020/2021 | |||||
|
H5Nx |
H5N1 | H5N3 | H5N4 | H5N5 | H5N8 | ||||
| Albanie | Volaille | 08/06/2021 | 6 |
|
6 | ||||
| Allemagne* | Sauvage | 30/10/2020 | 1 332 |
41 |
80 | 38 | 14 | 25 | 1 134 |
| Volaille | 04/11/2020 | 235 |
5 |
6 | 2 | 222 | |||
| Autriche | Sauvage | 04/02/2021 | 30 |
7 |
3 | 20 | |||
| Belgique | Sauvage | 13/11/2020 | 30 |
17 |
13 | ||||
| Volaille | 25/11/2020 | 2 |
1 |
1 | |||||
| Bulgarie | Sauvage | 31/03/2021 | 2 |
0 |
2 | ||||
| Volaille | 03/02/2021 | 8 |
8 |
||||||
| Croatie | Sauvage | 08/03/2021 | 3 |
0 |
3 | ||||
| Volaille | 21/11/2020 | 1 |
0 |
1 | |||||
| Danemark | Sauvage | 05/11/2020 | 318 |
5 |
3 | 2 | 7 | 301 | |
| Volaille | 16/11/2020 | 13 |
0 |
13 | |||||
| Espagne | Sauvage | 27/11/2020 | 3 |
1 |
2 | ||||
| Estonie | Sauvage | 12/02/2021 | 34 |
2 |
6 | 26 | |||
| Volaille | 18/02/2021 | 2 |
0 |
2 | |||||
| Finlande | Sauvage | 21/01/2021 | 34 |
6 |
16 | 12 | |||
| Volaille | 10/02/2021 | 1 |
0 |
1 | |||||
| France** | Sauvage | 27/11/2020 | 22 |
0 |
1 | 1 | 20 | ||
| Volaille | 16/11/2020 | 492 |
39 |
453 | |||||
| Grèce | Sauvage | 01/04/2021 | 4 |
0 |
4 | ||||
| Hongrie | Sauvage | 06/01/2021 | 3 |
0 |
1 | 2 | |||
| Volaille | 06/01/2021 | 7 |
1 |
1 | 5 | ||||
| Irlande | Sauvage | 06/11/2020 | 27 |
0 |
1 | 26 | |||
| Volaille | 10/12/2020 | 1 |
0 |
1 | |||||
| Italie | Sauvage | 20/11/2020 | 19 |
7 |
5 | 7 | |||
| Volaille | 23/01/2021 | 3 |
0 |
3 | |||||
| Kosovo | Volaille | 01/06/2021 | 7 |
0 |
7 | ||||
| Lettonie | Sauvage | 09/02/2021 | 37 |
0 |
1 | 36 | |||
| Lituanie | Sauvage | 08/01/2021 | 27 |
2 |
25 | ||||
| Volaille | 08/01/2021 | 39 |
0 |
39 | |||||
| Norvège | Sauvage | 27/11/2020 | 36 |
1 |
35 | ||||
| Pays-bas | Sauvage | 20/10/2020 | 114 |
8 |
34 | 1 | 2 | 1 | 68 |
| Volaille | 29/10/2020 | 12 |
3 |
9 | |||||
| Pologne | Sauvage | 09/12/2020 | 94 |
0 |
1 | 1 | 92 | ||
| Volaille | 24/11/2020 | 357 |
0 |
357 | |||||
| Roumanie | Sauvage | 09/01/2021 | 8 |
5 |
3 | ||||
| Volaille | 14/01/2021 | 8 |
0 |
1 | 7 | ||||
| Royaume-Uni*** | Sauvage | 08/11/2020 | 317 |
1 |
14 | 1 | 6 | 295 | |
| Volaille | 02/11/2020 | 21 |
0 |
2 | 19 | ||||
| Russie**** | Sauvage | 17/12/2020 | 7 |
|
1 | 4 | |||
| Volaille | 21/09/2020 | 19 |
|
18 | |||||
| Serbie | Sauvage | 06/03/2021 | 3 |
3 |
|||||
| Slovaquie | Sauvage | 07/01/2021 | 6 |
2 |
3 | 1 | |||
| Volaille | 22/01/2021 | 2 |
1 |
1 | |||||
| Slovénie | Sauvage | 23/11/2020 | 6 |
0 |
1 | 5 | |||
| Suède | Sauvage | 26/11/2020 | 109 |
17 |
1 | 1 | 18 | 72 | |
| Volaille | 16/11/2020 | 16 |
1 |
2 | 13 | ||||
| Suisse | Sauvage | 04/02/2021 | 2 |
1 |
1 | ||||
| Tchèque (Rép.) | Sauvage | 21/01/2021 | 28 |
2 |
1 | 25 | |||
| Volaille | 22/01/2021 | 37 |
1 |
36 | |||||
| Ukraine | Sauvage | 01/03/2021 | 4 |
4 |
|||||
| Volaille | 04/12/2020 | 14 |
10 |
4 | |||||
| Total Europe | Sauvage | 2 659 |
132 |
162 | 44 | 18 | 73 | 2 228 | |
| Volaille | 1 303 |
70 |
10 | 0 | 0 | 6 | 1 216 | ||
* dont un cas mixte H5N1/H5N8 (Allemagne, oiseaux sauvages)
** Données consolidées par le LNR
*** Source : OIE-WAHIS, DEFRA, correspondant à un décompte d’individus confirmés
**** Source : OIE-WAHIS
L’analyse des variations du nombre de cas et foyers hebdomadaires indique une épizootie en plusieurs temps, avec deux pics mi-novembre et mi-mars chez l’avifaune sauvage, et deux pics début janvier et autour d'avril chez les volailles (Figure 3 b). Pour l’avifaune sauvage, les dates pourraient correspondre à deux pics d’activité migratoire (une descendante, vers le sud, et une prénuptiale, vers le nord) avec des regroupements d’individus sur certaines haltes.
En faune sauvage, les pics de cas signalent une pression d’infection importante et donc une augmentation du risque d’introduction et de diffusion de virus IAHP dans les élevages les plus exposés. Le risque d’infection est en particulier accru dans les élevages de palmipèdes domestiques (espèces réceptives à l'infection par les virus IAHP circulant), les élevages sur parcours plein air non ou mal protégés des contacts avec l’avifaune sauvage, ou les élevages présentant des failles dans la mise en place ou l'observance des mesures de biosécurité (EFSA 2017).
Chez les volailles, les deux pics d’incidence observés, sont postérieurs à ceux observés dans le compartiment sauvage (deux mois pour les premiers pics, et moins d’un mois pour les seconds). Le pic du premier trimestre 2021 est dû à l’épizootie localisée dans le sud-ouest de la France, majoritairement dans des élevages de palmipèdes, et celui du second trimestre 2021, plus étalé, est dû à une forte propagation dans le compartiment domestique en Pologne, en particulier dans les élevages de type commercial, où les filières de production de Gallus, de canards et de dindes ont été touchées (Figures 3a, 4 et 5, EFSA Avian influenza overview December 2020 – February 2021, Avian influenza overview February – May 2021).
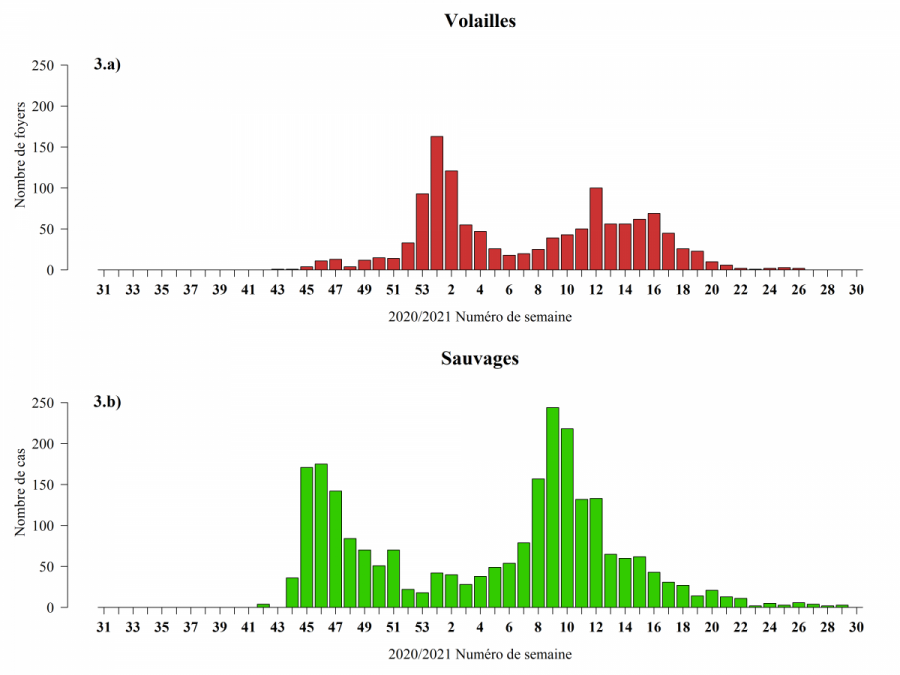
Figure 3 : Nombre de foyers (a) ou cas (b) d’IAHP H5 par compartiment en Europe et par semaine de suspicion ou de début d’évènement (ou date de confirmation en cas de donnée manquante) entre le 01/08/2020 et le 31/07/2021 inclus (Sources : Commission européenne ADNS, ADIS pour l’Europe, Empres-i pour l’Albanie, le Royaume-Uni et la partie Européenne de la Russie).
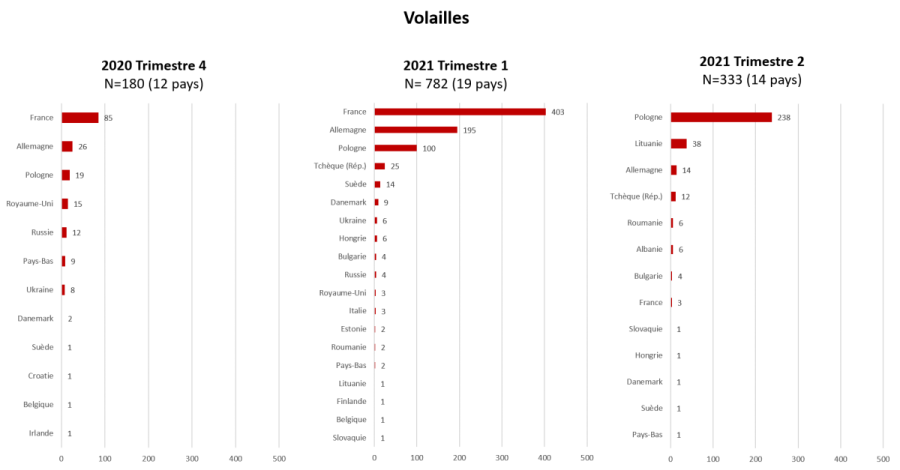
Figure 4 : Nombre de foyers domestiques d’IAHP H5 en Europe par pays (date de début d’évènement= suspicion ou date de confirmation en cas de donnée manquante) pour le 4ème trimestre 2020 et les 1er et 2nd trimestres 2021 (source : Commission européenne ADNS, ADIS et Empres-i).
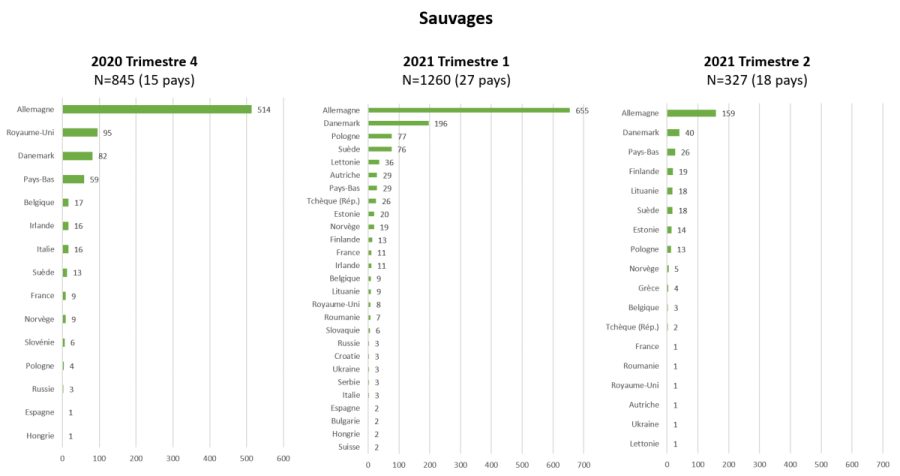
Figure 5 : Nombre de cas en faune sauvage d’IAHP H5 en Europe par pays (date de début d’évènement ou date de confirmation en cas de donnée manquante) pour le 4ème trimestre 2020 et les 1er et 2nd trimestres 2021 (source : Commission européenne ADNS, ADIS et Empres-i ).
A la fin du mois d’avril 2021, 95 % des foyers domestiques et 94 % des cas en faune sauvage de la période avaient été observés. Quelques foyers domestiques et cas en faune sauvage sporadiques ont été notifiés en mai, juin et juillet 2021. Le dernier foyer domestique en élevage professionnel a été observé le 04/07/2021 au Danemark, soit 35 semaines après le premier foyer observé sur la saison 2020-2021. En France, le dernier foyer domestique en basse-cour a été observé le 09/07/2021.
Le dernier cas sauvage a été observé le 30/07/2021 en Finlande, soit la dernière semaine de la période prise en compte dans cette note (source : Commission Européenne ADIS le 02/08/2021). Des foyers sporadiques chez les volailles et des cas en avifaune sauvage sont toujours observés au début du mois d’août 2021, en particulier en Pologne pour le compartiment domestique, et dans le nord de l’Europe dans les zones proches des sites de reproduction des espèces paléarctiques, pour le compartiment sauvage (source : Commission européenne ADIS le 13/08/2021). La figure 6 montre la répartition de l’ensemble des foyers et cas ayant débuté en Europe occidentale sur toute la période étudiée dans cette note, du 01/08/2020 au 31/07/2021.
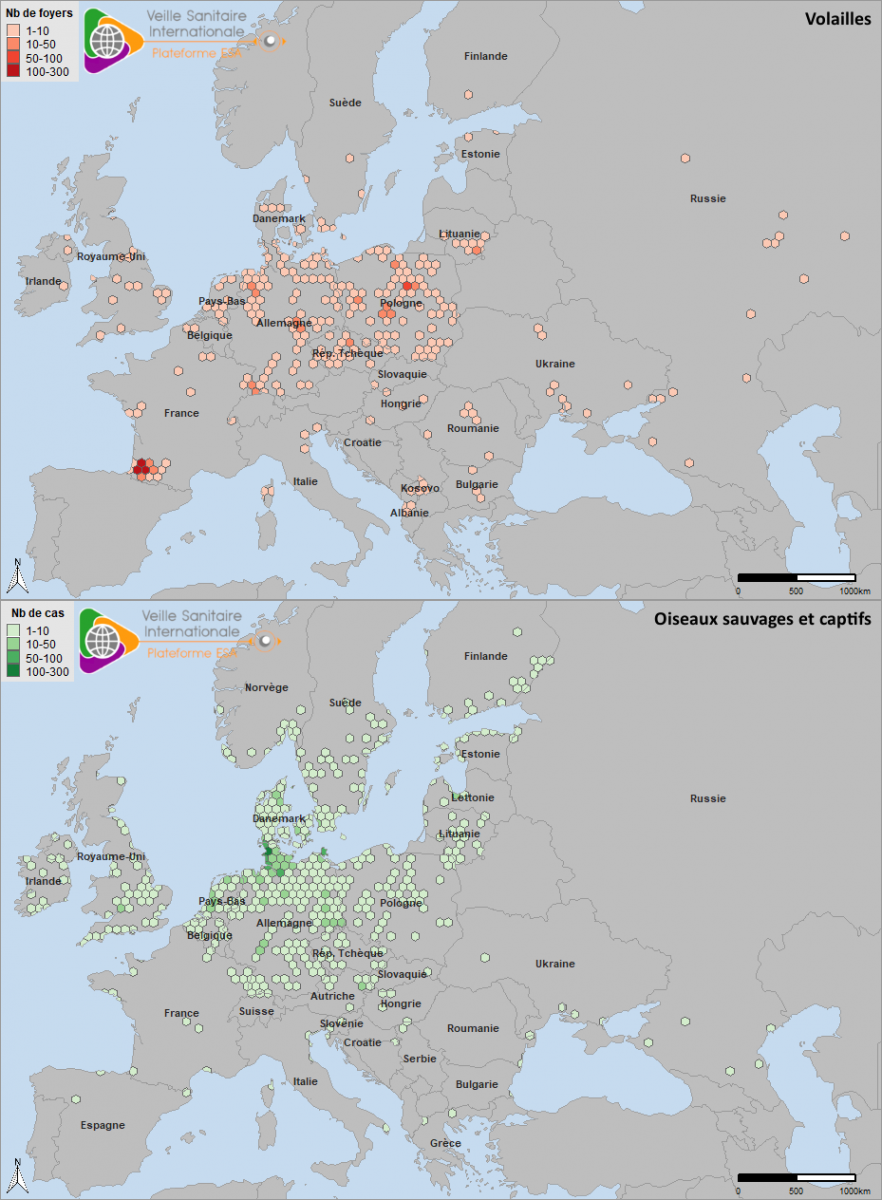
Figure 6. Densité des foyers domestiques (haut) et cas en avifaune sauvage (bas) d’IAHP en Europe occidentale, ayant débuté entre le 01/08/2020 et le 31/07/2021 (source : Commission européenne ADNS, ADIS, OIE).
En France, les premiers foyers domestiques ont été identifiés en Corse (n=7) à partir du 16/11/2020, en lien épidémiologique avec un autre foyer détecté dans une animalerie des Yvelines. L’historique et la localisation des foyers détectés et investigués en Haute-Corse et Corse-du-Sud sont précisés dans le BHVSI-SA du 15/12/2020. Le neuvième foyer a été confirmé le 06/12/2020 dans les Landes. Puis le virus s'est propagé rapidement, dans le Sud-Ouest de la France, avec des détections dans les Landes, les Hautes-Pyrénées, les Pyrénées-Atlantiques, le Lot-et-Garonne et le Gers. Le 12/12/2020, un foyer a été détecté en Vendée, puis le 13/12/2020 dans le département limitrophe des Deux-Sèvres. Au total, 492 foyers domestiques (475 dans le Sud-Ouest et 17 hors Sud-Ouest) ont été déclarés cette saison.
Vingt-deux cas ont été confirmés en avifaune (autres que volailles), dont un cas sur des oiseaux sauvages libres en lien épidémiologique avec un foyer en élevage et deux cas concernant des oiseaux captifs. Sept oiseaux sauvages infectés H5N8 auraient été contaminés secondairement, à partir de foyers en élevages de volailles : en Corse (2), dans les Landes (1), dans les Ardennes (3) et dans le Haut-Rhin (1) (sources: ANSES avis le 26/05/2021, BHVSI le 22/06/2021). Trois sous-types différents de virus IAHP, H5N8, H5N3 et H5N1, ont été caractérisés (Tableaux 1 et 2). Un épisode de mortalité massive a été observé dans le département de la Manche le 21/01/2021, où des bécasseaux maubèches (Calidris canutus) ont été retrouvés morts sur une plage (au moins 300 oiseaux morts sur un groupe de 10 000 bécasseaux maubèches). Les quatre cas confirmés (sur quatre testés) étaient associés à une infection par un virus d’IAHP H5N3 (sources : LNR, OFB).
Origine du virus et aspects génétiques
Une première analyse phylogénétique (Lewis et al. 2021) à partir des séquences des virus d’IAHP H5N8 détectés entre fin juillet et début novembre 2020 en Russie, au Kazakhstan et dans les trois premiers États membres de l’UE touchés (Pays-Bas, Allemagne, Royaume-Uni) chez des poulets, des oies, des canards domestiques et chez plusieurs espèces d’oiseaux sauvages (Anatidés et rapaces) a confirmé que tous ces virus appartenaient au clade 2.3.4.4b. Ils étaient tous directement apparentés au virus IAHP H5N8 détecté en Irak en mai 2020 et apparentés à un même génotype ancestral précédemment détecté en 2016-2018, lors de la précédente vague épizootique intercontinentale qui a concerné l’Asie, l’Europe et l’Afrique : il s’agissait du génotype majoritaire identifié alors en Europe par Lycett et al. 2020. Ces virus n’étaient pas apparentés aux virus d’IAHP H5N8 de clade 2.3.4.4b qui avaient circulé en Allemagne et en Europe de l’Est de décembre 2019 à juin 2020. Cette analyse suggère également que ces mêmes virus ont pu persister et circuler depuis 2018, en l’absence de toute détection, au sein d’élevages de volailles galliformes sans que l’étendue et la localisation de cette circulation puissent être précisées.
Les parentés phylogénétiques décrites dans cette première étude sont confirmées par Beerens et al. (2021), qui ont plus précisément décrit et daté l’émergence d’un virus H5N8 identifié aux Pays-Bas chez deux cygnes tuberculés trouvés morts le 17/10/2020. Leur analyse suggère qu’un ancêtre de ce virus possédant une composition génomique similaire aurait pu circuler à partir de mars 2017 et être responsable de foyers d’IAHP en Égypte en 2018-2019 (Beerens et al. 2021).
L’analyse phylogénétique des génomes complets de virus IAHP H5 identifiés entre juillet 2020 et mai 2021, provenant de vingt États membres de l’UE, du Royaume-Uni, de la Norvège, de la Suisse, du Kazakhstan et de la Russie, montre que (Source : EFSA scientific report le 31/05/2021 et LRUE Standing Committee on Plants, Animals, Food and Feed (PAFF) 17/06/2021) (cf. encadré 1):
- l’ensemble des séquences H5 forme un groupe homogène unique appartenant au clade 2.3.4.4b de la lignée A/goose/Guandong/1/1996,
- ces virus ont probablement circulé de Sibérie occidentale vers le nord de l’Europe, puis vers le sud et l’est de l’Europe, avec transmission possible en Afrique de l’Ouest (LRUE PAFF 17/06/2021),
- au moins dix-sept génotypes distincts circulent conjointement en Asie Centrale et en Europe, résultant de différents événements de réassortiment avec des virus IA faiblement ou hautement pathogènes précédemment détectés chez les oiseaux sauvages ou domestiques.
Huit génotypes associés au sous-type H5N5, six génotypes pour H5N8, un génotype pour H5N1, un génotype pour H5N3 et un génotype pour le sous-type H5N4 des virus IAHP circulants ont été caractérisés au cours de la saison 2020-2021 en Europe (Figure 7). La majorité des foyers et cas détectés chez les oiseaux est associée à un génotype dominant de sous-type H5N8 (génotype A, figure 7) : ce génotype a également été associé à plusieurs cas d’infection chez des mammifères sauvages en Europe. Ainsi, au Royaume-Uni, l’infection par ce génotype de virus IAHP H5N8 avait été détectée chez trois phoques (phoque gris Halichoerus grypus et phoque commun Phoca vitulina) et un renard roux (Vulpes vulpes) hébergés dans un centre de sauvegarde et de réhabilitation de la faune sauvage où ceux-ci ont probablement été contaminés par des cygnes tuberculés qui avaient séjourné dans le même centre et avaient succombé à l’infection par le même virus (Floyd et al. 2021). Ce même génotype a également été identifié chez un phoque gris en Suède. Sept cas d’infection rapportée comme inapparente chez l’Homme ont également été signalés en Russie (ville d’Astrakhan, décembre 2020) (WHO le 02/03/2021, Pyankova et al. 2021).
Enfin, la mise en évidence d’un virus d’IAHP H5N1, proche des virus ayant circulé chez les oiseaux sauvages en Europe, a été signalée chez deux renards aux Pays-Bas, province de Groningue (WUR le 28/05/2021).
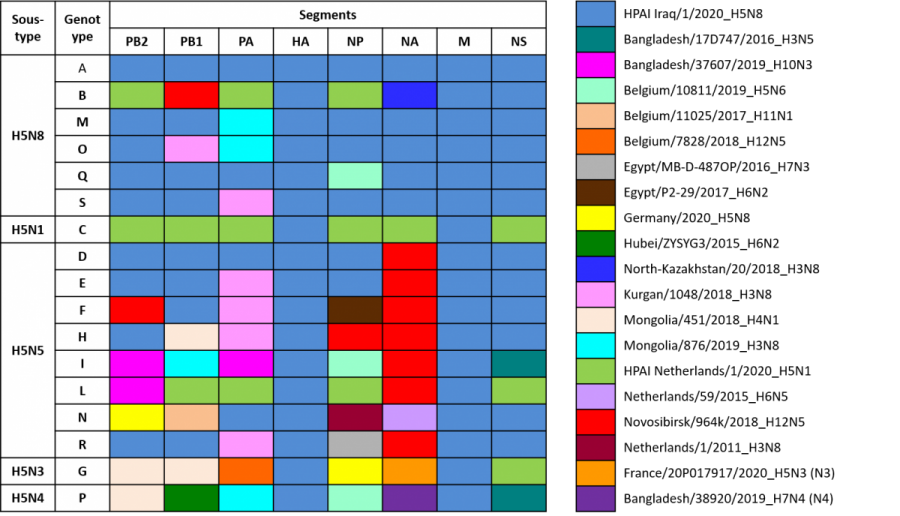
Figure 7. Représentation schématique des génotypes des virus IAHP H5 identifiés en Eurasie depuis mai 2020 (source: LRUE PAFF 17/06/2021). Pour chaque segment du génome viral et chaque groupe phylogénétique distinct identifié par une couleur différente, le nom abrégé d’un virus IA de référence est associé dans la légende (cette nomenclature est uniquement descriptive et ne représente pas nécessairement la séquence ancestrale du groupe phylogénétique).
Les analyses des séquences de virus IAHP H5 identifiées chez les oiseaux (et réalisées par le LRUE) ont montré la présence sporadique et isolée de mutations associées avec une adaptation du virus aux mammifères ou avec une augmentation de sa virulence chez ces derniers : y compris certaines mutations majeures, également rapportées chez les virus identifiés lors des infections chez les mammifères au Royaume-Uni et en Suède. Toutefois, en l’absence d’autres mutations adaptatives ou compensatoires venant les renforcer, ces mutations encore isolées n’augmenteraient pas de façon significative le risque de transmission du virus des oiseaux aux mammifères ou entre mammifères, au-delà des cas de transmission sporadique aux mammifères déjà constatés au cours de cet épisode. L’analyse des génomes complets de virus IAHP H5N8 identifiés dans différents foyers en France, réalisée par le LNR Anses Ploufragan, conduit aux mêmes conclusions. Plusieurs marqueurs d’adaptation aux volailles galliformes terrestres et de virulence chez celles-ci ont également été détectés chez des virus H5N8 identifiés depuis janvier 2021 chez les oiseaux sauvages et les volailles, et chez un virus H5N3 prélevé chez un oiseau sauvage (Source : EFSA scientific report le 31/05/2021 ).
La Lituanie a confirmé le 26/03/2021 l’infection de deux cygnes tuberculés par un virus IAHP H7N7. Cette détection d’un virus IAHP de sous-type H7 dans l’avifaune sauvage était remarquable, un seul précédent ayant été déclaré en Algérie en octobre 2016 lors d’une mortalité massive d’oiseaux sauvages imputée à un virus IAHP H7N1.
Rôle des oiseaux sauvages migrateurs
Les oiseaux d'eau migrateurs, plus particulièrement les anatidés (canards, oies, cygnes et espèces apparentées), peuvent jouer différents rôles dans l’épidémiologie du virus : révélateur ou sentinelles, réservoir, ou propagateurs. De 2005 à 2020, des virus IAHP H5 ont été détectés chez au moins 86 espèces d'oiseaux échantillonnées en Europe. La majorité des virus IAHP H5 ont été isolés à partir d'oiseaux trouvés morts, mais ont pu également être isolés à partir d'oiseaux apparemment sains de plusieurs espèces : cela a par ailleurs été rapporté au cours de la saison 2020-2021, chez trois espèces d’Anatidés, canard siffleur (M. penelope), canard colvert (Anas platyrhynchos) et sarcelle d’hiver (Anas crecca). La présence du virus de l'IAHP H5 et/ou des anticorps spécifiques de l'IAHP H5 chez les oiseaux vivants suggère que certains individus de ces espèces d'oiseaux peuvent de manière asymptomatique être porteurs du virus et participer à la propagation du virus sur de longues distances au cours des migrations (Verhagen, Fouchier, et Lewis 2021). La voie d'introduction la plus probable des virus de l'IA dans l'UE sont les routes migratoires Nord-Est et Est (source : OFB, EFSA Panel on Animal Health and Welfare (AHAW) 2017).
Sur la saison 2020-2021, 2 430 notifications en faune sauvage ou captive ont été faites sur ADNS ou ADIS, d’au moins 20 familles d’espèces d’oiseaux, dont 78 %[3] sont inféodées au milieu aquatique (Figure 8). Les oies et les cygnes totalisent la majorité des déclarations, ce qui est cohérent avec les observations de mortalité massive et étendue rapportées sur les côtes de la mer du Nord et de la Baltique. Cette saison est également marquée par des épisodes plus localisés de mortalité importante de limicoles (bécasseaux maubèches le plus souvent) associés au virus IAHP H5N3 dans plusieurs pays (source : LNR).
Plusieurs cas concernent des oiseaux non migrateurs (moineaux, buses variables) qui laissent à penser que le virus aurait ensuite diffusé du compartiment domestique aux oiseaux commensaux des élevages de volailles et pourrait laisser craindre l’installation durable du virus et de la maladie (sources: ANSES avis le 26/05/2021,BHVSI le 22/06/2021, OFB).
*dont Bernaches
** Rapaces nocturnes et diurnes
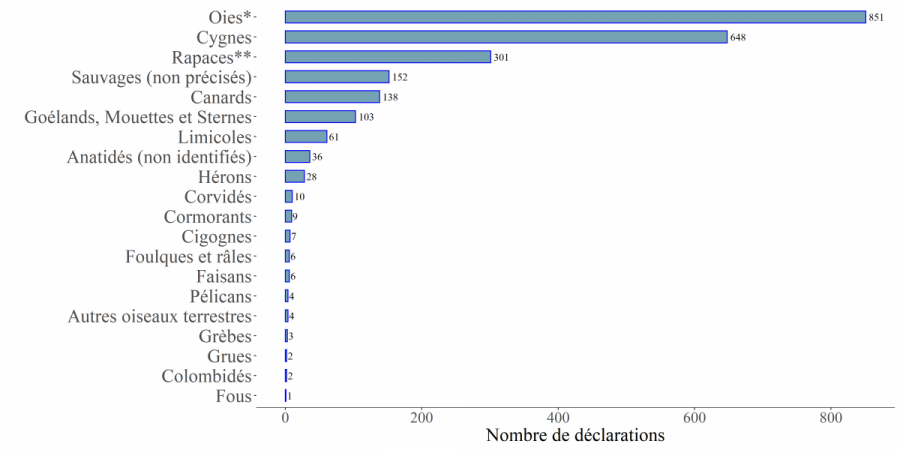
Figure 8. Nombre de déclarations relatives aux cas en faune sauvage d'IAHP en Europe sur ADNS-ADIS du 01/08/2020 au 31/07/2021 par famille ou groupe d’espèces d’oiseaux. ATTENTION : Le nombre de déclarations ne reflète pas le nombre de spécimens concernés (il peut y avoir plusieurs oiseaux pour une seule déclaration) (Source : Commission européenne ADIS au 01/08/2021).
En France, les virus IAHP ont été introduits en France continentale lors de la migration post-nuptiale «descendante » de fin d’automne (source : ANSES avis le 26/05/2021). La présence de virus IAHP a été confirmée sur douze espèces d’oiseaux sauvages, appartenant à quatre familles (Tableau 2). La liste chronologique des cas sauvages identifiés en France est disponible en Annexe du BHVSI du 06/07/2021 .
Tableau 2 : Sous-type de virus IAHP identifiés en France du 01/08/2020 au 31/07/2021 par espèce d’oiseau sauvage, et par ordre de fréquence du sous-type.
|
Sous-type
|
Espèce d’oiseau sauvage
|
|
|
H5N8
|
Bernache cravant (Branta bernicla)
Oie cendrée (Anser anser) Tadorne de Belon (Tadorna tadorna) Cygne tuberculé (Cygnus olor) Mouette tridactyle (Rissa tridactyla) Goéland leucophée (Larus michahellis) Moineau domestique (Passer domesticus) Buse variable (Buteo buteo) Cigogne blanche (Ciconia ciconia) Faucon pélerin (Falco peregrinus) |
|
|
H5N3
|
Bécasseau maubèche (Calidris canutus)
Courlis cendré (Numenius arquata) |
|
|
H5N1
|
Bernache cravant (Branta bernicla)
|
|
|
H5Nx
|
Pigeon ramier (Columba palumbus)
|
La saison 2020-2021 au regard des précédentes saisons
Le nombre de foyers domestiques d’IAHP en Europe pour la saison 2020-2021 (n=1 303) est plus important que les précédentes saisons (figure 9). Il équivaut à celui observé en 2016-2017 (n=1 136) (source : Commission européenne ADIS du 01/08/2016 au 31/07/2017). La distribution géographique des foyers présente également des similitudes avec la saison 2016-2017, caractérisée par une épizootie très localisée dans le Sud-Ouest de la France et diffuse sur l’Est de l’Europe (figure 10). La saison est marquée par une extension vers le nord de l’Europe, alors que le virus IAHP avait circulé l’année précédente principalement dans la partie Est de l’Europe. Des foyers domestiques ont été déclarés pour la première fois en Estonie, Finlande, Irlande, Kosovo, Lituanie et Norvège.
La saison 2020-2021 est marquée par un nombre de déclarations sans précédent de cas en faune sauvage (n= 2 659), soit +51 % par rapport à 2016-2017 (n= 1 614) (source : Commission européenne ADIS du 01/08/2016 au 31/07/2017). Cette augmentation dans le compartiment sauvage est à mettre en lien avec les épisodes de forte mortalité observés dans l’avifaune en Allemagne, au Danemark et aux Pays-Bas, et avec l’augmentation des efforts de surveillance, en particulier de la surveillance programmée en 2020 en Allemagne et Belgique, et la concentration des prélèvements collectés dans le cadre de la surveillance évènementielle au quatrième trimestre 2020 en France, Danemark, Luxembourg et Norvège. Au total en 2020, 18 968 oiseaux sauvages ont été collectés (12 418 par surveillance évènementielle et 6 550 par surveillance programmée) contre 8 934 en 2019, 9 145 en 2018 et 19 543 en 2017 (source : EFSA rapport annuel de surveillance 2020). L’épizootie 2020-2021 a une durée supérieure à celle des années précédentes et la décrue est plus tardive, touchant les poussins en avifaune sauvage.
La distribution géographique des cas dans l’avifaune sauvage est marquée par un nombre élevé de déclarations dans le nord de l’Europe, en particulier dans les pays scandinaves et sur la péninsule du Jutland (nord Allemagne et Danemark). Des cas dans l’avifaune sauvage ont été déclarés pour la première fois en Estonie, Lettonie et en Norvège (Madslien et al. 2021).
Cette distribution diffère de la saison 2016-2017, qui avait débuté avec un important épicentre autour du lac de Constance, touchant la Suisse et les pays voisins. Au cours de l’épizootie 2016-2017, les détections s’étaient ensuite largement étendues vers le Nord et l’Est de l’Europe, jusqu’aux Balkans (figure 11).
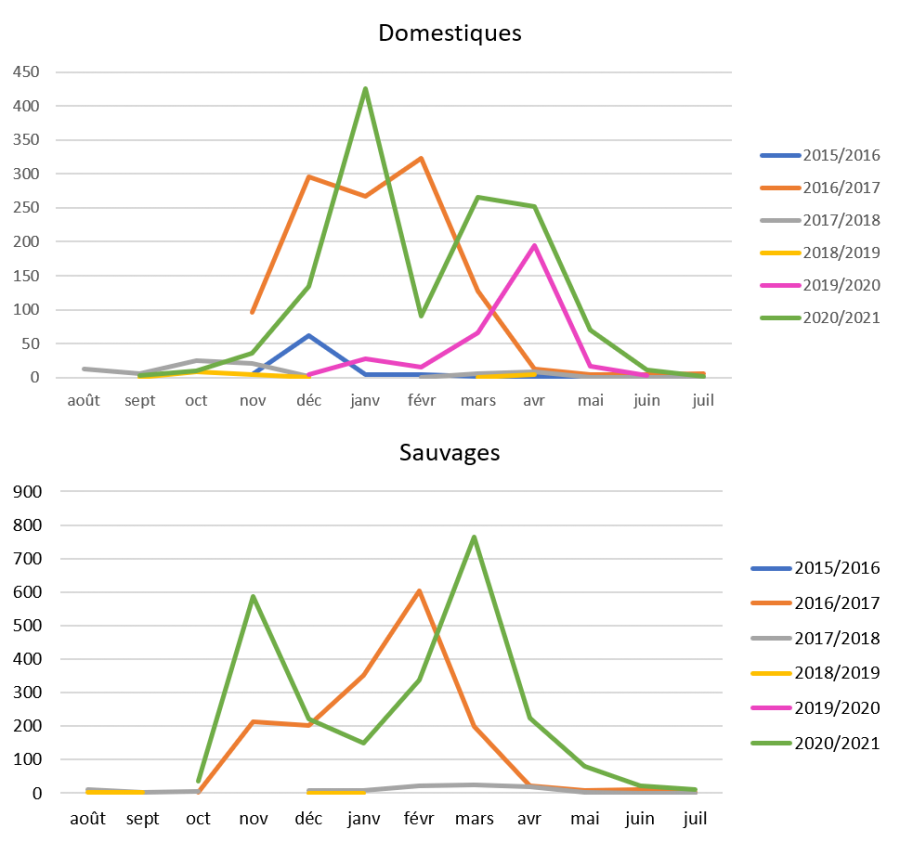
Figure 9 : Nombre de cas en faune sauvage (en bas) et de foyers domestiques en Europe (en haut) par année et par mois de début d’évènement pour les saisons 2015-2016, 2016-2017, 2017-2018, 2018-2019, 2019-2020 et 2020-2021 (source : Commission Européenne ADIS). ATTENTION : échelles différentes entre les deux graphes.
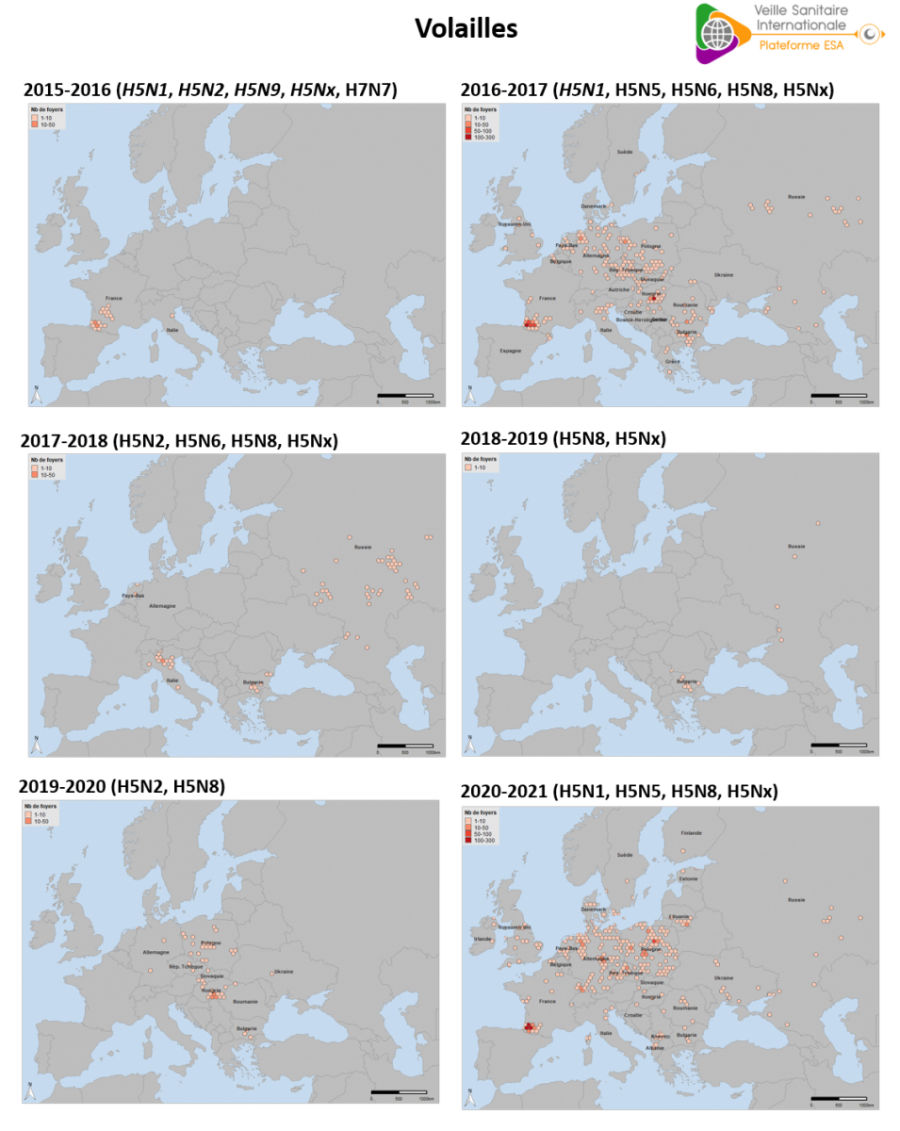
Figure 10. Densité des foyers domestiques d’IAHP en Europe occidentale, par saison de 2015-2016 à 2020-2021 (chaque saison étant bornée du 01/08 de l’année n-1 au 31/07 de l’année n). Les différents virus IAHP de sous-type H5 mentionnés ici appartiennent tous au clade 2.3.4.4b de la lignée A/goose/Guandong/1/1996, à l’exception de l’ensemble des virus IAHP de sous-type H5 détectés en 2015-2016 et du virus de sous-type H5N1 détecté en 2016-2017 (en italique) (source : Commission européenne ADNS, ADIS, OIE).
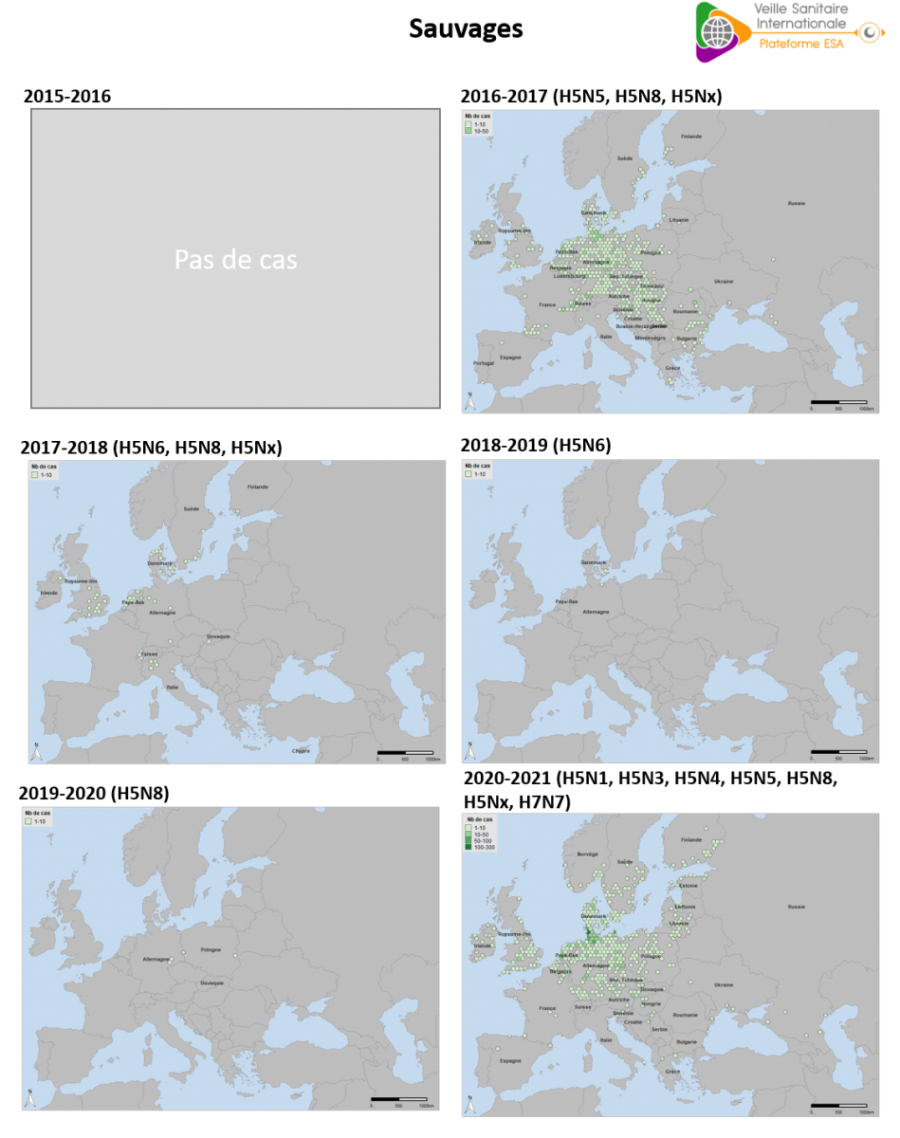
Figure 11. Densité de cas dans l’avifaune sauvage d’IAHP en Europe occidentale, par saison 2015-2016 à 2020-2021 (chaque saison étant bornée du 01/08 de l’année n-1 au 31/07 de l’année n). Les différents virus IAHP de sous-type H5 mentionnés ici appartiennent tous au clade 2.3.4.4b de la lignée A/goose/Guandong/1/1996 (source : Commission européenne ADNS, ADIS, OIE).
| Pour en savoir plus |
|---|
|
Références bibliographiques
Beerens, Nancy, Rene Heutink, Frank Harders, Marit Roose, Sylvia B. E. Pritz-Verschuren, Evelien A. Germeraad, et Marc Engelsma. 2021. « Incursion of Novel Highly Pathogenic Avian Influenza A(H5N8) Virus, the Netherlands, October 2020 - Volume 27, Number 6—June 2021 - Emerging Infectious Diseases Journal - CDC ». https://doi.org/10.3201/eid2706.204464.
EFSA. 2017. « Avian Influenza ». EFSA Journal 15 (10): e04991. https://doi.org/10.2903/j.efsa.2017.4991.
Floyd, Tobias, Ashley C. Banyard, Fabian Z. X. Lean, Alexander M. P. Byrne, Edward Fullick, Elliot Whittard, Benjamin C. Mollett, et al. 2021. « Systemic Infection with Highly Pathogenic H5N8 of Avian Origin Produces Encephalitis and Mortality in Wild Mammals at a UK Rehabilitation Centre ». https://doi.org/10.1101/2021.05.26.445666.
Lewis, Nicola S., Ashley C. Banyard, Elliot Whittard, Talgat Karibayev, Thamer Al Kafagi, Ilya Chvala, Alex Byrne, et al. 2021. « Emergence and spread of novel H5N8, H5N5 and H5N1 clade 2.3.4.4 highly pathogenic avian influenza in 2020 ». Emerging Microbes & Infections 10 (1): 148‑51. https://doi.org/10.1080/22221751.2021.1872355.
Lycett, Samantha J., Anne Pohlmann, Christoph Staubach, Valentina Caliendo, Mark Woolhouse, Martin Beer, Thijs Kuiken, et Global Consortium for H5N8 and Related Influenza Viruses. 2020. « Genesis and Spread of Multiple Reassortants during the 2016/2017 H5 Avian Influenza Epidemic in Eurasia ». Proceedings of the National Academy of Sciences, août. https://doi.org/10.1073/pnas.2001813117.
Madslien, Knut, Torfinn Moldal, Britt Gjerset, Sveinn Gudmundsson, Arne Follestad, Elliot Whittard, Ole-Herman Tronerud, et al. 2021. « First detection of highly pathogenic avian influenza virus in Norway ». BMC Veterinary Research 17 (1): 218. https://doi.org/10.1186/s12917-021-02928-4.
Pyankova, Olga G., Ivan M. Susloparov, Anastasia A. Moiseeva, Natalia P. Kolosova, Galina S. Onkhonova, Aleksey V. Danilenko, Elena V. Vakalova, et al. 2021. « Isolation of Clade 2.3.4.4b A(H5N8), a Highly Pathogenic Avian Influenza Virus, from a Worker during an Outbreak on a Poultry Farm, Russia, December 2020 ». Eurosurveillance 26 (24): 2100439. https://doi.org/10.2807/1560-7917.ES.2021.26.24.2100439.
Verhagen, Josanne H., Ron A. M. Fouchier, et Nicola Lewis. 2021. « Highly Pathogenic Avian Influenza Viruses at the Wild–Domestic Bird Interface in Europe: Future Directions for Research and Surveillance ». Viruses 13 (2): 212. https://doi.org/10.3390/v13020212.
[1] Le Kazakhstan a notifié en septembre et octobre 2020 un foyer dans l’avifaune sauvage et plusieurs foyers domestiques quasi-exclusivement dans la région Nord du pays (source : OIE au 16/09 et 05/10/2020). Huit régions administratives (soit quatre de plus que celles des déclarations officielles) ont été concernées par des détections en élevage (sources : Promed le 26/09/2020, le 06/10/2020 et article de presse du 28/09/2020).
[2] Israël a notifié plusieurs foyers et cas au sein de l’avifaune captive d’IAHP entre le 13 et le 19/10/2020. Il est indiqué que la source probable de l’infection serait une contamination via des oiseaux sauvages migrateurs (source : notification OIE du 13/10/2020 et rapport de suivi du 19/10/2020).
[3] Anatidés (Oies, Cygnes et Canards), Laridés (Goélands, Mouettes et Sternes), Podicipedidés (Grèbes), Ciconidés (Cigognes), Ardeidés (Hérons), Pélicanidés (Pélicans), Phalacrocoracidés (Cormorans), Sulidés (Fous), Rallidés (Foulques et Râles), Gruidés (Grues), Scolopacidés, Charadriidés et Haematopodidés (Limicoles), Busard des roseaux (Circus aeruginosus) et Pygargue à queue blanche (Haliaeetus albicilla).